This web page was produced as an assignment for an undergraduate course at Davidson College
Introduction:
This page aims to give a brief evolutionary history of the sodium-potassium pump by taking a look at orthologs of this protein in other organisms. At its most basic level, the sodium-potassium pump, aka the Na,K-ATPase, evolved as an adaptation to handle osmotic burdens on cells with semi-permeable membranes. Other functions and the basic structure of the protein can be reviewed by clicking this link. Cells contain material which cannot diffuse through its own membrane such as nucleic acids, proteins, and metabolites; however, most cells are surrounded by fluid and ions that are more or less able to flow freely into and out of the cell. Why is this a problem? It's a problem because both the cytoplasm of the cell and the external medium will attempt to achieve equilibrium, leaving the cell vulnerable to flooding or shriveling depending on the relative ionic concentration of the external medium. Plants and bacteria handled this osmotic problem by evolving a cellulose wall to prevent the diffusion of ions across their membranes. However, this adaptation did not come without a catch as these cells are condemned to a sessile existence, restrained by the very adaptation that keeps them alive. The sodium-potassium pump has been proclaimed to be the the enzyme responsible for the metazoan's ability to both move without the resistance of a cellulose wall and to maintain an osmotic balance. The function of the sodium-potassium pump has already been discussed in some detail on the webpage which can be reached by clicking on the link given above, so I will not delve into it on this page. It will suffice to say that the pump compensates by actively pumping ions across the cell membrane to effectively parry any potentially fatal osmotic pressures (Stein, 1995).
Ortholog Comparison:
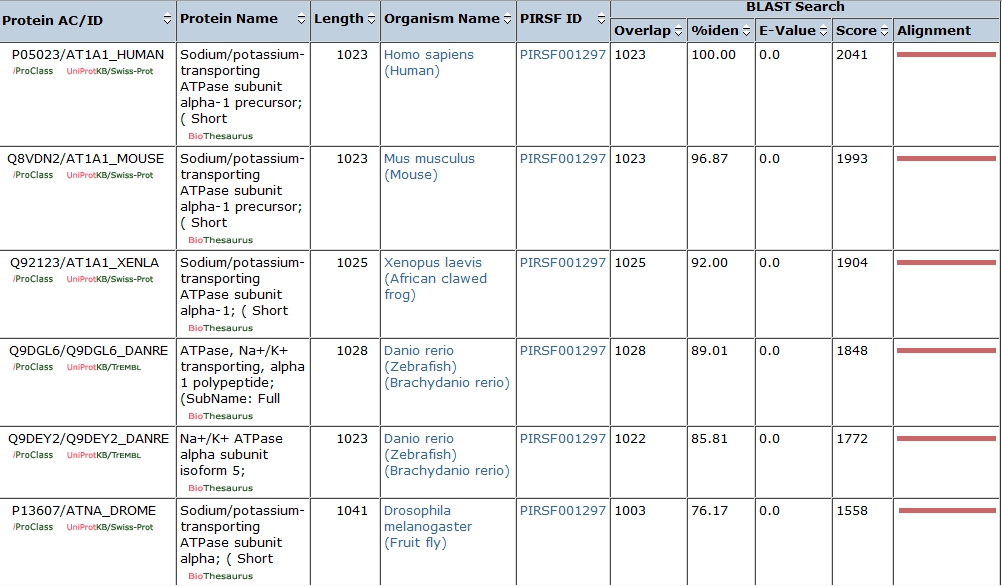
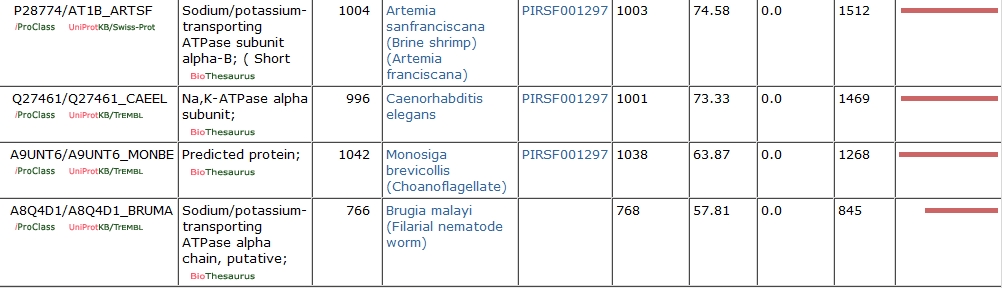
Using the Protein Information Resource (http://pir.georgetown.edu/), I searched for orthologs of the amino acid sequence of the human alpha 1 isoform of the sodium-potassium pump. Using several tools available on this site, I was able to compare the sequences of the orthologs I found. The following table lists all of the orthologs I searched for, including statistical analysis of the conformity between the proteins.
I picked these because the alpha-N,K-ATPase subunit is a metazoan adaptation, and I felt that these species were sufficient to tell the evolutionary story from choanoflagellates through humans.
The first broad comparison I performed was a multiple alignment of the ortholog sequences. This is a very long list of each sequence stacked on top of the other, with dashes, dots, and asterisks at the bottom indicating how well conserved each amino acid is.Since the alignment takes up a lot of space, I will show only a few sections which show the most conservation between the different sequences analyzed (Figure 1).
Figure 1

These highly conserved sequences are, presumably, directly involved in the specific catalytic functions of the alpha-subunit of the sodium-potassium pump. The differences in the highly conserved sequences (mainly occurring in the nematodes (Q27461,A8Q4D1), the choanoflagellate (A9UNT6), and at times in the brine shrimp (P28774)) could be due to adaptations of these organisms to their particular environments. These adaptations may change the enzyme's sodium or potassium affinity to best suit the salinity of its environment. In the case of the brine shrimp, the differences may be a result of the accidental inclusion of a different alpha subunit isoform. I believe I may have included the brine shrimp's alpha-B isoform, as opposed to its alpha-1 isoform. THE CONCEPT OF ISOFORMS WILL BE DISCUSSED LATER.
Another useful graphic the multiple alignment tool gave me was a tree that showed the relatedness of the sequences in a different manner.
Figure 2
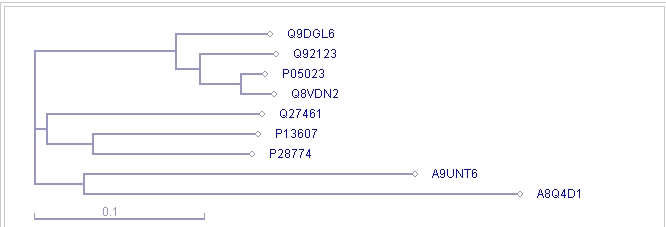
Once again, please refer to the key to distinguish between the species analyzed. Notice the closeness of the two mammals, the mouse, Mus musculus,(Q8VDN2) and the human, Homo sapien, (P05023), in the tree. Also note, the relatedness between the amphibian (Q92123) and the mammalian mouse and human subgroup. Finally, at the very top of the tree we have a zebrafish, Danio rerio,(Q9DGL6). The branches of the top four sequences fits nicely into the vertebrate evolutionary story of fish to amphibian to mammal. The next three sequences, according to this tree, also share a common ancestral protein. The nematode,C. elegans (Q27461), the fruit fly, Drosophila melanogaster (P13607), and the brine shrimp, Artemia Franciscana, (P28774) are all invertebrates and possess closely related alpha-1 subunit protein sequences. Also fairly close to the aforementioned group is the choanoflagellate, Monosiga brevicollis (A9UNT6), and the falarial nematode, Brugia malayi (A8Q4D1). Of particular interest to us is the choanoflagellate because it is a living relative of a pre-metazoan ancestor. Therefore, it could help us determine the origin of the alpha-1 sodium-potassium pump subunit. The closeness of the nematodes with the choanoflagellate makes perfect sense given the fact that the the choanoflagellate is a sister group of the metazoans and the nematode represents an early metazoan (Saez et. al., 2009). This tree suggests that two closely related alpha-1 subunits evolved from the choanoflagellate in nematodes and insects, but did not persist in the animal groups at the top of the tree. Another alpha-1 subunit, perhaps originally a paralog of the nematode/insect alpha-1 subunit, seems to have persisted in the higher animal groups at the top of the table.
A tool aptly called domain display analyzed each sequence for known sections of the sequences known to be associated with the certain enzyme functions. An analysis of the ortholog sequences I found shows conserved domains associated with cation transporting ATPases, which is precisely what the sodium-potassium pump is (Figure 3).
Figure 3

The key is included in the figure.
Notice that all except for the filarial nematode possess all four domains typical of the family of P-type ATPases, which includes the alpha subunit of the Na, K-ATPase.
Brief Summary of a Useful Review Article:
This data, which I collected myself using the aforementioned website and the tools available on it, corresponds fairly well with the work done in a review article called Evolutionary history of Na,K-ATPAses and their osmoregulatory role by Saez et. al.. In this article, they take a look at both metazoans and non-metazoans in an effort to achieve a big evolutionary picture of the alpha-subunit from its ancestral proteins through to the bonafide Na,K-ATPase alpha subunit proteins. Figure 4, borrowed from the Saez paper, shows a tree of non-metazoan organisms possessing P-type ATPase genes.
Figure 4
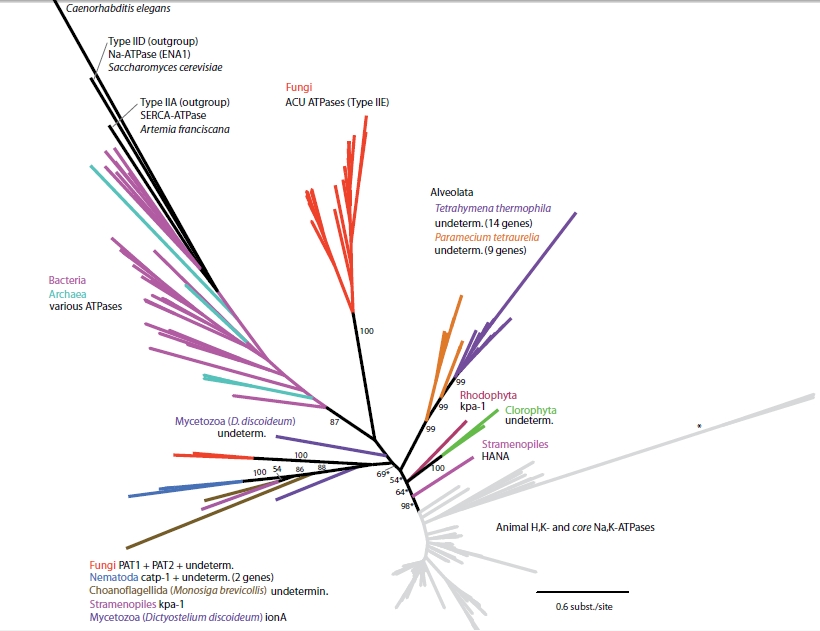
Within the P-type family, there are a number of subfamilies. The Na,K-ATPase alpha subunit belongs to the P-type ATPase IIC subfamily, whereas other P-type ATPases fall into IIA, IIB, IID, etc. By looking at this tree, the writers first inferred that the IIC subfamily probably originated within the prokaryotes because the outgroup sequences are all within the prokaryotic "clad" of this tree. They then point out the fungal proteins closer to the animal "core" proteins, called PAT1 and PAT2. Evidence has been presented that these two proteins are actually Na,K-ATPases, which would place the ancestral N,K-ATPase before the evolutionary split between fungi and metazoa (Saez et. al, 2009). The authors also comment on the proximity of the choanoflagellate and nematoda, proteins; the significance of which I explained earlier using the sequences I analyzed. The next figure (Figure 5)represents an enlarged version of the bottom part of the tree in the previous figure. This portion of the tree contains organisms who possess isoforms of the real IIC Na,K-ATPase.
Figure 5
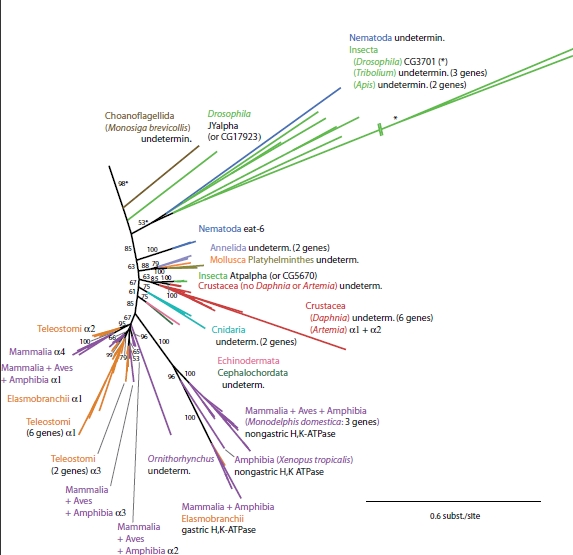
It is important to notice the one M. brevicollis IIC ATPase located at the base of this tree. The writers suggest that this choanoflagellate actually made two P-type ATPases; one of which became the alpha-Na,K-ATPases found in the nematodes, and the other evolved into the majority of the current animal Na,K-ATPases. Support is given for this idea by the confirmation of a single amino acid difference(asparagine to glutamine) at a particular site in this M. brevicollis ATPase which makes the Na,K-ATPase resistant to the drug ouabain. This fits in nicely with the preexisting idea that ancestral Na,K-ATPase proteins are resistant to this drug. They also confirm that the amino acid segments responsible for alpha-beta complex formation are present in the protein.
The paper also examines the evolutionary significance of an organism possessing more than one isoform of the alpha-Na,K-ATPase. For example, organisms which live in environments with fluctuating osmotic pressures may evolved multiple isoforms to be expressed under different conditions within their environment. For example, the brine shrimp, A. franciscana, has an alpha-2 isoform which features two lysine amino acids in positions where asparagine residues typically reside in the alpha-1 sequence. The positive charge of the lysines actually gives the enzyme the ability to operate under stonger osmotic pressures by reducing the number of ions transported per each catalytic cycle. Mammals actually possess four different isoforms which exhibit different expression patterns and functions.
References:
Saez AG, Lozano E, Zaldivar-Riveron A. Evolutionary history of Na,K-ATPases and their osmoregulatory role. Genetica 2009; 136(3): 479-490
Stein WD. The sodium pump in the evolution of animal cells. Biological Sciences 1995; 1329(349); 263-269
Go back Home
Go to the Molecular Web Page
Contact me at: ropalmer@davidson.edu